- 上一篇:水杨酸对镉胁迫下一年生黑麦草的生长生理影响
- 下一篇:霍乱弧菌基因组中非编码DNA功能的初步研究
2.4 酵母双杂交筛选Dsl1复合体亚基与Ⅲ型PI3K复合体亚基之间的互作关系 13
3 讨论 14
致谢 14
参考文献: 15
附录一:测序结果及序列比对 16
附录二:培养基及配方 22
附录三:试剂与缓冲液配方 23
筛选酿酒酵母中与Dsl1复合体亚基互作的细胞自噬相关蛋白
引言:在真核生物中,细胞自噬(Autophagy)是在进化中高度保守的能调控细胞内物质周转的重要降解过程,且它是能依赖于细胞内的溶酶体或液泡(酵母)介导的胞内物质降解途径。细胞自噬普遍存在于真核生物中,是细胞在自噬相关基因的调控下将自身体内一些损坏的蛋白或细胞器等通过自噬途径降解用于新物质的合成,使胞内物质得以循环利用[1],进而影响生物体的生理过程。如植物中细胞自噬参与生长、发育、衰老、免疫反应以及对事物和非生物胁迫等的应答过程[2-4];动物中细胞自噬参与免疫反应、肿瘤形成、神经退行性疾病及衰老等过程[5-7]。
细胞自噬现象在真核生物中广泛存在,由于其在细胞内的重要作用,酿酒酵母则是研究细胞自噬分子机制最为充分的模式生物。迄今酵母菌中已发现41个基因直接参与细胞自噬过程,并被命名为细胞自噬相关基因(AuTophagy-related Genes,ATG)[7]。在细胞自噬发生的过程中,除上述自噬基因所编码的自噬蛋白参与调控外,细胞内一些参与其它生理过程的蛋白,尤其是囊泡运输相关蛋白在细胞自噬过程中的作用也不断有报道[8-11]。
在真核细胞中,与细胞分泌和内吞途径的膜融合有关的细胞器之间的交流需要囊泡运输来介导。运输囊泡在形成后可以在细胞骨架上扩散或被积极地转运至受体细胞器上。为了使囊泡能够运输到正确的目标受体膜上,在囊泡与受体室的膜融合之前需先进行校正,该过程叫做栓留。在栓留期间,受体室上的栓留因子与囊泡相互作用,推测认为囊泡可能与该细胞器相互识别,然后使囊泡更加接近受体膜。当栓留将囊泡紧贴到受体膜之后,发生被称为锚定的第二次校正,启动SNARE蛋白(Soluble NSF Attachment Protein Receptor)的配对。目标受体膜上的SNARE蛋白能够识别运输囊泡上的SNARE蛋白,这两种蛋白在成功识别之后,会形成SNARE蛋白复合体,以使囊泡和受体膜紧密接触[12],最后在SM蛋白(Sec1/Munc18蛋白)等因子的作用下发生膜的融合[13](图1-1)。
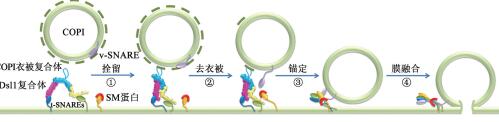
Dsl1复合体是种拴系因子,由三种组分组成:Dsl1、Dsl3和Tip20,该复合体是目前发现的最简单的大分子多亚基复合体,位于内质网上,用于启动由高尔基体产生的COPI囊泡和内质网膜的融合[12]。DSL1和DSL3被鉴定为SLY1(一种在内质网与高尔基体之间穿梭的t-SNARE相互作用蛋白)的遗传相互作用因子,TIP20首先是在内质网SNARE SEC20的研究中被发现的。Dsl1复合体的亚基Dsl3、Tip20可以分别通过与内质网膜上的SNARE蛋白的结合而使其牢固的绑定到内质网膜上[15](图1-2),并且它们之间的结合能够调节由高尔基体产生的COPI囊泡与内质网的融合。研究表明,Dsl1复合体是一个塔状结构,Dsl3和Tip20亚基一起形成塔的基部,其底部包含有两个与内质网上的SNARE蛋白的结合位点,并在其顶端提供了一个用于捕获COPI囊泡的位于Dsl1亚基中间附近的柔性套索[16]。Dsl1复合体通过其N末端的调节结构域与内质网膜上的SNARE蛋白结合,并与组装的SNARE复合体结合;而且,它还能够促进SNARE复合体的组装[16](图1-2)。综上所述,Dsl1复合体能够在细胞质中捕获COPI(coat protein I)囊泡,然后将其拉近至内质网膜,最后将膜结合在一起进行融合。