- 上一篇:蜜蜂螺原体黏附相关蛋白对宿主意蜂免疫的影响
- 下一篇:解淀粉芽孢杆菌SQR9根际互作微生物的初步鉴定
转座子是基因组的重要组成部分,越来越多的转座子在动植物、真菌、细菌等的不同物种中被鉴定出来,在基因组较大(>2000 Mb)的被子植物中,转座子可占基因组的50%以上[7],在基因组较小的生物如水稻(430 Mb)中,转座子占的比例约为35%[8]。
80年代初期,随着植物中第一个转座子的成功克隆,转座子研究进入了分子遗传学的领域。随着新一代测序技术和生物信息技术的发展,转座子的种类、数量和功能被大尺度地发掘,关于转座子起源、特性、调控等方面的研究取得了颇多进展,人们对转座子的认识越来越深入。尽管转座子的起源和存在意义一直备受争议,但越来越多证据表明转座子影响着基因组的大小和组成、基因的表达与调控等众多方面。根据转座机制,转座子可以分为两类。I型转座子(反转录转座子)是利用“复制-粘贴”的方式,即转座子DNA先转录为RNA,然后以转录的RNA为中间媒介,将它作为模板,被逆转录酶反转录成DNA,最后将dsDNA在整合酶或核酸内切酶的作用下插入宿主基因组中[9]。II型转座子(DNA转座子)则是用“剪切-粘贴”的方式,即DNA在转座酶的作用下从染色体中分离出来,再整合到基因组其他位置[9]。从转座机制来看,反转录转座子还可以分为LTR反转录转座子(主要包括gypsy 和copia两大类)和non-LTR反转录转座子(LTR:长末端重复,Long terminal repeat; non-LTR:非长末端重复,Non-long terminal repeat)。如图1所示[10]:(a)许多DNA转座子两翼有末端反向重复(terminal inverted repeats ,TIRs;黑色箭头 ),中间序列编码转座酶(紫色圆圈)。转座时,转座酶结合或接近TIRs,将转座子从原位点上剪切下来,再整合至新的位点上。由于靶位点两条链的切开位点是交错的,因此整合后两侧会形成靶位点重复(TSD,target-site duplication )(b) LTR反转录转座子包含两个长末端重复序列(LTRs,黑色箭头)和编码对逆转录及转座至关重要的蛋白Gag、蛋白酶、逆转录酶和整合酶。5'LTR含有由宿主RNA聚合酶II识别的启动子序列,产生TE的mRNA(直角箭头表示转录起始点)。随后,将TE mRNA(浅蓝色波浪线)、转录酶(粉色圆圈)和整合酶被组装到由Gag蛋白(小粉红色圆圈)形成的病毒样颗粒中,逆转录酶将TE mRNA逆转录成全长dsDNA。接着,整合酶(紫色圆圈)将cDNA(宽的深蓝色弧)插入新的目标序列中。 类似于DNA转座子的转座酶,反转录转座子整合酶可以产生交错切割位点,产生TSDs。(c)非LTR反转录转座子缺乏LTRs,有一个或两个ORF。与LTR反转录转座子不同,非LTR反转录转座子的转录是产生全长的mRNA(浅蓝色波浪线)。接下来,元件编码的内切核酸酶在靶位点DNA中产生单链切口,释放出3'-OH用于引发mRNA的逆转录,这种机制被称为通过靶位点引发的逆转录(TPRT)。由自主非LTR反转录转座子编码的蛋白质还可以动员非自主反转录转座子的RNA以及其他胞内RNA。图中描绘了长散布元件1(L1)的TPRT机制,一些非LTR反转录转座子缺乏3'poly(A)尾。非LTR反转录转座子的整合可导致TSDs或者在基因组DNA靶位点上有小量缺失。
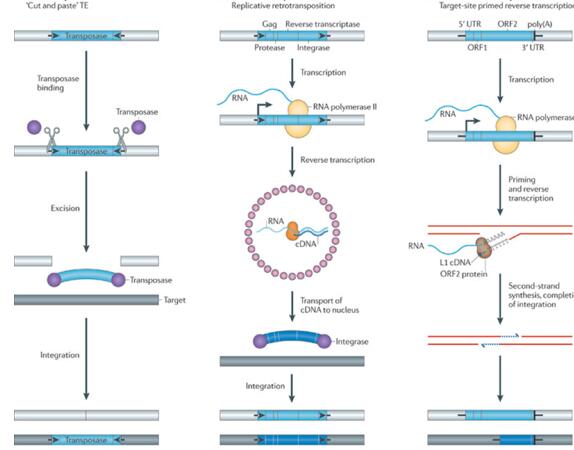
图1三种常见的植物转座机制[10]
Fig.1 The perse mechanisms of transposon mobilization[10]
根据转座子的编码情况,上述两类转座子都可以再分为自主型转座子和非自主型转座子。自主型转座子可以编码转座所需的蛋白,非自主型转座子则不能,并需要依赖同类的自主型转座子进行转座。但是之前的I型转座子和II型转座子的分类方式不完全适用所有转座子,比如在细菌和真菌中发现了不以RNA为中间媒介的“复制-粘贴”型转座。因此,为解决这些冲突,目前普遍采用是否以RNA为中间媒介(class)、DNA转座过程中是否被复制(subclass)、插入机制(order)、蛋白保守性和靶位点重复序列即TSD序列长度(superfamily)、DNA序列尤其是转座酶序列的保守性(family)、自主和非自主(subfamily)这种综合考虑的分级分类方式(表1)[11]。在水稻中,转座子分布如图(表2)[12],常见分类的亚家族有DNA转座子:Transposase_24 亚家族、 Ac/Ds亚家族、 Mariner亚家族、 Mutator亚家族、Pong亚家族、 CACTA亚家族, En/Spm亚家族、 MuDR亚家族;逆转录转座子: SINE亚家族、LINE亚家族、centromere-specific亚家族、Ty1-copia亚家族、 Ty3-gypsy亚家族。